Genetik sürüklenme
| Biyoloji alt dalı |
| Evrimsel biyoloji |
|---|
 |
| Ana maddeler |
| Tarihçe Genel bakış Rönesans ve Aydınlanma Darwin Öncesi Darwin • Türlerin Kökeni Sentezden Önce Modern evrimsel sentez Moleküler evrim • Evo-devo Güncel Araştırmalar |
| Doğa tarihi Yaşam tarihi • Evrimsel hayat ağacı Biyoçeşitlilik • Biyocoğrafya Sınıflandırma • Kladistik Paleontoloji Filogenetik |
| Süreç ve sonuçlar Popülasyon genetiği Genetik çeşitlilik Mutasyon Doğal seçilim • Adaptasyon Polimorfizm Genetik sürüklenme • Gen akışı Türleşme • Adaptif radyasyon Yardımlaşma • Birlikte evrim Soy tükenmesi |
| Sosyal çıkarımlar |
| Diğer alanlar |
| Evrimsel biyoloji portalı Kategori • İlgili başlıklar • Kitap |
|
Rastgele genetik sürüklenme, alel sürüklenmesi veya Wright etkisi[1] olarak da bilinen genetik sürüklenme, bir popülasyondaki mevcut bir gen varyantının (alel) frekansında rastgele şansa bağlı olarak meydana gelen değişimdir.[2]
Genetik sürüklenme, gen varyantlarının tamamen yok olmasına ve böylece genetik çeşitliliğin azalmasına neden olabilir.[3] Ayrıca başlangıçta nadir olan alellerin çok daha sık görülmesine ve hatta sabitlenmesine de neden olabilir.
Bir alelin az sayıda kopyası mevcut olduğunda, genetik sürüklenmenin etkisi daha belirgindir ve çok sayıda kopya mevcut olduğunda, etki daha az belirgindir (büyük sayılar yasası nedeniyle). 20. yüzyılın ortalarında, genetik sürüklenme de dahil olmak üzere doğal seçilime karşı nötr süreçlerin göreceli önemi üzerine şiddetli tartışmalar yaşanmıştır. Mendel genetiğini kullanarak doğal seçilimi açıklayan Ronald Fisher,[4] genetik sürüklenmenin evrimde en fazla küçük bir rol oynadığı görüşünü savunmuş ve bu görüş birkaç on yıl boyunca baskın görüş olarak kalmıştır. 1968'de popülasyon genetikçisi Motoo Kimura, genetik bir değişikliğin bir popülasyonda yayıldığı çoğu durumun (fenotiplerde değişiklik olması gerekmese de) nötr mutasyonlar üzerinde etkili olan genetik sürüklenmeden kaynaklandığını iddia eden moleküler evrimin nötral teorisiyle tartışmayı yeniden alevlendirdi.[5][6] 1990'larda, karmaşık sistemlerin nötral geçişler yoluyla nasıl ortaya çıktığını açıklamaya çalışan yapıcı nötral evrim önerilmiştir.[7][8]
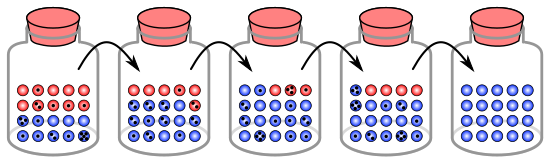
Kavanozdaki bilyeler analojisi
Genetik sürüklenme süreci, bir popülasyondaki 20 organizmayı temsil etmek üzere bir kavanozdaki 20 bilye kullanılarak gösterilebilir.[9] Bu bilye kavanozunu başlangıç popülasyonu olarak düşünün. Kavanozdaki bilyelerin yarısı kırmızı, yarısı mavidir ve her bir renk popülasyondaki bir genin farklı bir aleline karşılık gelir. Her yeni nesilde organizmalar rastgele çoğalır. Bu üremeyi temsil etmek için, orijinal kavanozdan rastgele bir bilye seçin ve aynı renkte yeni bir bilyeyi yeni bir kavanoza koyun. Bu, orijinal bilyenin "yavrusudur", yani orijinal bilye kavanozunda kalır. Bu işlemi ikinci kavanozda 20 yeni bilye olana kadar tekrarlayın. İkinci kavanoz artık 20 "yavru" ya da çeşitli renklerde bilye içerecektir. İkinci kavanoz tam olarak 10 kırmızı ve 10 mavi bilye içermediği sürece, alel frekanslarında rastgele bir değişim meydana gelmiştir.
Bu işlem birkaç kez tekrarlanırsa, her nesilde seçilen kırmızı ve mavi bilye sayıları dalgalanır. Bazen bir kavanozda "ebeveyn" kavanozdan daha fazla kırmızı bilye, bazen de daha fazla mavi bilye bulunur. Bu dalgalanma genetik sürüklenmeye benzer - bir nesilden diğerine alellerin dağılımındaki rastgele bir varyasyondan kaynaklanan popülasyonun alel frekansındaki bir değişiklik.
Herhangi bir nesilde, belirli bir renkte hiçbir bilye seçilmemiş olabilir, yani hiçbir yavruları yoktur. Bu örnekte, hiç kırmızı bilye seçilmezse, yeni nesli temsil eden kavanoz sadece mavi yavrular içerir. Bu durumda, kırmızı alel popülasyonda kalıcı olarak kaybolurken, kalan mavi alel sabitlenmiş olur: gelecekteki tüm nesiller tamamen mavidir. Küçük popülasyonlarda sabitlenme sadece birkaç nesilde gerçekleşebilir.
Olasılık ve alel frekansı
Genetik sürüklenme mekanizmaları basitleştirilmiş bir örnekle açıklanabilir. Bir damla çözelti içinde izole edilmiş çok büyük bir bakteri kolonisi düşünün. Bakteriler, A ve B olarak etiketlenmiş iki alele sahip tek bir gen dışında genetik olarak özdeştir; bunlar nötral alellerdir, yani bakterilerin hayatta kalma ve üreme yeteneklerini etkilemezler; bu kolonideki tüm bakterilerin hayatta kalma ve üreme olasılığı eşittir. Bakterilerin yarısının A aleline, diğer yarısının da B aleline sahip olduğunu varsayalım. Böylece, A ve B'nin her biri 1/2 alel frekansına sahiptir.
Daha sonra çözelti damlası, yalnızca dört bakterinin yaşamasına yetecek kadar yiyeceğe sahip olana kadar küçülür. Diğer tüm bakteriler üremeden ölür. Hayatta kalan dört bakteri arasında, A ve B alelleri için 16 olası kombinasyon mevcuttur:
(A-A-A-A), (B-A-A-A), (A-B-A-A), (B-B-A-A),
(A-A-B-A), (B-A-B-A), (A-B-B-A), (B-B-B-A),
(A-A-A-B), (B-A-A-B), (A-B-A-B), (B-B-A-B),
(A-A-B-B), (B-A-B-B), (A-B-B-B), (B-B-B-B).
Orijinal çözeltideki tüm bakterilerin çözelti küçüldüğünde hayatta kalma olasılığı eşit olduğundan, hayatta kalan dört bakteri orijinal koloniden rastgele bir örneklemdir. Hayatta kalan dört kişinin her birinin belirli bir alele sahip olma olasılığı 1/2'dir ve bu nedenle çözelti küçüldüğünde herhangi bir alel kombinasyonunun ortaya çıkma olasılığı:

(Orijinal popülasyon boyutu o kadar büyüktür ki örnekleme etkin bir şekilde değiştirme ile gerçekleşir). Başka bir deyişle, 16 olası alel kombinasyonunun her birinin 1/16 olasılıkla gerçekleşme olasılığı eşittir.
Aynı sayıda A ve B içeren kombinasyonlar sayıldığında aşağıdaki tablo elde edilir:
| A | B | Kombinasyonlar | Olasılık |
| 4 | 0 | 1 | 1/16 |
| 3 | 1 | 4 | 4/16 |
| 2 | 2 | 6 | 6/16 |
| 1 | 3 | 4 | 4/16 |
| 0 | 4 | 1 | 1/16 |
Tabloda gösterildiği gibi, B aleli ile aynı sayıda A aleline sahip kombinasyonların toplam sayısı altıdır ve bu kombinasyonun olasılığı 6/16'dır. Diğer kombinasyonların toplam sayısı ondur, dolayısıyla eşit olmayan sayıda A ve B aleli olasılığı 10/16'dır. Dolayısıyla, orijinal koloni eşit sayıda A ve B aleli ile başlamış olsa da, büyük olasılıkla, kalan dört üyeli popülasyondaki alel sayısı eşit olmayacaktır. Eşit sayı durumu aslında eşit olmayan sayı durumundan daha az olasıdır. İkinci durumda, popülasyonun alel frekansları rastgele örnekleme nedeniyle değiştiği için genetik sürüklenme meydana gelmiştir. Bu örnekte popülasyon, popülasyon darboğazı olarak bilinen bir olgu ile sadece dört rastgele hayatta kalana kadar daralmıştır.
Hayatta kalan A (veya B) aleli kopyalarının sayısı için olasılıklar (yukarıdaki tablonun son sütununda verilmiştir) doğrudan binom dağılımından hesaplanabilir; burada "başarı" olasılığı (belirli bir alelin mevcut olma olasılığı) 1/2'dir (yani, kombinasyonda k adet A (veya B) aleli kopyası olma olasılığı) ile verilir:

Burada n=4 hayatta kalan bakteri sayısıdır.
Matematiksel modeller
Genetik sürüklenmenin matematiksel modelleri ya dallanma süreçleri ya da idealize edilmiş bir popülasyonda alel frekansındaki değişiklikleri tanımlayan bir difüzyon denklemi kullanılarak tasarlanabilir.[10]
Wright-Fisher modeli
A veya B olmak üzere iki aleli olan bir gen düşünün. Diploidide, N bireyden oluşan popülasyonlar her genin 2N kopyasına sahiptir. Bir birey aynı alelin iki kopyasına veya iki farklı alele sahip olabilir. Wright-Fisher modeli (adını Sewall Wright ve Ronald Fisher'dan almıştır) nesillerin üst üste gelmediğini (örneğin, yıllık bitkilerin yılda tam olarak bir nesli vardır) ve yeni nesilde bulunan genin her kopyasının eski nesildeki genin tüm kopyalarından bağımsız olarak rastgele çekildiğini varsayar. Son nesilde p frekansına sahip bir alelin k kopyasını elde etme olasılığını hesaplamak için formül şu şekildedir:[11][12]

burada "!" sembolü faktöriyel fonksiyonunu göstermektedir. Bu ifade binom katsayısı kullanılarak da formüle edilebilir,

Moran modeli örtüşen nesiller varsayar. Her zaman adımında, bir birey üremek ve bir birey de ölmek üzere seçilir. Dolayısıyla her zaman adımında, belirli bir alelin kopya sayısı bir artabilir, bir azalabilir ya da aynı kalabilir. Bu da stokastik matrisin üç köşeli olduğu anlamına gelir ki bu da Moran modeli için matematiksel çözümlerin Wright-Fisher modeline göre daha kolay olduğu anlamına gelir. Öte yandan, Wright-Fisher modeli kullanılarak bilgisayar simülasyonlarının gerçekleştirilmesi genellikle daha kolaydır, çünkü daha az zaman adımının hesaplanması gerekir. Moran modelinde, bir nesilden geçmek için N zaman adımı gerekir, burada N etkin popülasyon boyutudur. Wright-Fisher modelinde ise bu süre sadece birdir.[13]
Uygulamada, Moran ve Wright-Fisher modelleri niteliksel olarak benzer sonuçlar verir, ancak genetik sürüklenme Moran modelinde iki kat daha hızlı çalışır.
Diğer sürüklenme modelleri
Eğer yavru sayısındaki varyans Wright-Fisher modeli tarafından varsayılan binom dağılımının verdiğinden çok daha büyükse, o zaman aynı genel genetik sürüklenme hızı (varyans etkili popülasyon büyüklüğü) göz önüne alındığında, genetik sürüklenme seçilime kıyasla daha az güçlü bir kuvvettir.[14] Aynı varyans için bile, yavru sayısı dağılımının yüksek momentleri binom dağılımınınkileri aşarsa, genetik sürüklenmenin gücü yine önemli ölçüde zayıflar.[15]
Örnekleme hatası dışındaki rastgele etkiler
Alel frekanslarındaki rastgele değişiklikler, örnekleme hatası dışındaki etkilerden de kaynaklanabilir, örneğin seçilim baskısındaki rastgele değişiklikler gibi.[16]
Belki de genetik sürüklenmeden daha önemli olan önemli bir alternatif stokastiklik kaynağı genetik otostoptur.[17] Genetik çekim, bağlantılı lokuslar üzerindeki seçilimin bir lokus üzerindeki etkisidir. Genetik çekimin matematiksel özellikleri genetik sürüklenmeden farklıdır.[18] Alel frekansındaki rastgele değişimin yönü, nesiller arasında otokorelasyon gösterir.[2]
Sürüklenme ve sabitlenme
Hardy-Weinberg ilkesi, yeterince büyük popülasyonlarda denge; göç, genetik mutasyonlar veya seçilim nedeniyle bozulmadığı sürece alel frekanslarının bir nesilden diğerine sabit kaldığını belirtir.[19]
Bununla birlikte, sonlu popülasyonlarda, bir sonraki nesle aktarılan alellerin rastgele örneklenmesinden yeni aleller kazanılmaz, ancak örnekleme mevcut bir alelin kaybolmasına neden olabilir. Rastgele örnekleme bir aleli ortadan kaldırabildiğinden ancak yerine yenisini koyamadığından ve alel frekansındaki rastgele düşüşler veya artışlar bir sonraki nesil için beklenen alel dağılımlarını etkilediğinden, genetik sürüklenme bir popülasyonu zaman içinde genetik tekdüzeliğe doğru yönlendirir. Bir alel 1 (%100) frekansına ulaştığında popülasyonda "sabitlendiği" söylenir ve bir alel 0 (%0) frekansına ulaştığında kaybolur. Daha küçük popülasyonlar fiksasyona daha hızlı ulaşırken, sonsuz popülasyon sınırında fiksasyona ulaşılamaz. Bir alel sabitlendiğinde, genetik sürüklenme durur ve mutasyon veya gen akışı yoluyla popülasyona yeni bir alel eklenmedikçe alel frekansı değişemez. Dolayısıyla, genetik sürüklenme rastgele ve yönsüz bir süreç olsa da, zaman içinde genetik varyasyon ortadan kaldıracak şekilde hareket eder.[20]
Sürüklenme nedeniyle alel frekansı değişim oranı

Genetik sürüklenmenin bir alel üzerinde etkili olan tek evrimsel güç olduğu varsayıldığında, p ve q alel frekansları ile başlayan birçok çoğaltılmış popülasyonda t nesil sonra, bu popülasyonlar boyunca alel frekansındaki varyans şöyledir:

Sabitlenme veya kayba kadar geçen süre
Genetik sürüklenmenin bir alel üzerinde etkili olan tek evrimsel güç olduğunu varsayarsak, herhangi bir zamanda bir alelin popülasyonda sabitlenme olasılığı basitçe o anda popülasyondaki frekansıdır.[22] Örneğin, A aleli için p frekansı %75 ve B aleli için q frekansı %25 ise, sınırsız zaman verildiğinde A'nın popülasyonda sabitlenme olasılığı %75 ve B'nin sabitlenme olasılığı %25'tir.
Sabitlenmenin gerçekleşmesi için beklenen nesil sayısı popülasyon büyüklüğü ile orantılıdır, öyle ki küçük popülasyonlarda fiksasyonun çok daha hızlı gerçekleşeceği tahmin edilmektedir.[23] Normalde bu olasılıkları belirlemek için toplam popülasyondan daha küçük olan etkin popülasyon büyüklüğü kullanılır. Etkin popülasyon (Ne), soy içi üreme düzeyi, popülasyonun en küçük olduğu yaşam döngüsü aşaması ve bazı nötr genlerin seçilim altında olan diğer genlerle genetik olarak bağlantılı olduğu gerçeği gibi faktörleri dikkate alır.[14] Etkin popülasyon büyüklüğü aynı popülasyondaki her gen için aynı olmayabilir.[24]
Wright-Fisher modeline göre, nötral bir alelin genetik sürüklenme yoluyla sabitlenmesinden önce beklenen süreyi yaklaşık olarak hesaplamak için kullanılan ileriye dönük bir formül şöyledir:

Burada T nesil sayısı, Ne etkin popülasyon büyüklüğü ve p verilen alel için başlangıç frekansıdır. Sonuç, belirli büyüklüğe (Ne) ve alel frekansına (p) sahip bir popülasyonda belirli bir alel için fiksasyon gerçekleşmeden önce geçmesi beklenen nesil sayısıdır.[25]
Nötral alelin genetik sürüklenme yoluyla kaybolması için beklenen süre şu şekilde hesaplanabilir:[11]

Bir mutasyon, başlangıç frekansının ihmal edilebilir olması için yeterince büyük bir popülasyonda yalnızca bir kez ortaya çıktığında, formüller şu şekilde basitleştirilebilir:[26]

Nötral bir mutasyonun sabitlenmesinden önce beklenen ortalama nesil sayısı için ve

Gerçek boyutu N olan bir popülasyonda nötr bir mutasyonun kaybından önce beklenen ortalama nesil sayısı için.[27]
Hem sürüklenme hem de mutasyon ile kayba kadar geçen süre
Yukarıdaki formüller, bir popülasyonda halihazırda mevcut olan ve ne mutasyona ne de doğal seçilime tabi olan bir alel için geçerlidir. Bir alel, mutasyonla kazanıldığından çok daha sık mutasyonla kaybediliyorsa mutasyon ve sürüklenme, kayba kadar geçen süreyi etkileyebilir. Mutasyonel kayba eğilimli alel popülasyonda sabit olarak başlarsa ve replikasyon başına m oranında mutasyonla kaybedilirse, haploid bir popülasyonda kaybına kadar nesiller içinde beklenen süre şu şekilde verilir:
![{\displaystyle {\bar {T}}_{\text{lost}}\approx {\begin{cases}{\dfrac {1}{m}},{\text{ if }}mN_{e}\ll 1\\[8pt]{\dfrac {\ln {(mN_{e})}+\gamma }{m}}{\text{ if }}mN_{e}\gg 1\end{cases}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/0dca9b7dc746a7b2a8d8a63770ac53782d1639e3)
Burada Euler sabitidir.[28] İlk yaklaşım, kayıp için belirlenen ilk mutanta kadar geçen bekleme süresini temsil eder; kayıp daha sonra genetik sürüklenme ile nispeten hızlı bir şekilde gerçekleşir ve 1/m ≫ Ne. zaman alır. İkinci yaklaşım, mutasyon birikimi yoluyla deterministik kayıp için gereken süreyi temsil eder. Her iki durumda da sabitlenmeye kadar geçen süre 1/m terimi aracılığıyla mutasyon tarafından domine edilir ve etkin popülasyon büyüklüğünden daha az etkilenir.

Doğal seçilime karşı genetik sürüklenme
Doğal popülasyonlarda, genetik sürüklenme ve doğal seçilim tek başına hareket etmez; her iki olgu da mutasyon ve göç ile birlikte her zaman iş başındadır. Nötral evrim, tek başına sürüklenmenin değil, hem mutasyonun hem de sürüklenmenin ürünüdür. Benzer şekilde, seçilim genetik sürüklenmeyi bastırdığında bile, yalnızca mutasyonun sağladığı varyasyon üzerinde etkili olabilir.
Doğal seçilimin, evrimi mevcut çevreye kalıtsal adaptasyonlara doğru yönlendiren bir yönü varken, genetik sürüklenmenin bir yönü yoktur ve sadece şans matematiği tarafından yönlendirilir.[29] Sonuç olarak, sürüklenme, fenotipik etkilerine bakılmaksızın bir popülasyon içindeki genotipik frekanslar üzerinde etkili olur. Buna karşılık seçilim, fenotipik etkileri taşıyıcılarının hayatta kalmasını ve/veya üremesini artıran alellerin yayılmasını destekler, olumsuz özelliklere neden olan alellerin frekanslarını düşürür ve nötral olanları göz ardı eder.[30]
Büyük sayılar yasası, alelin mutlak kopya sayısı az olduğunda (örneğin, küçük popülasyonlarda), nesil başına alel frekansları üzerindeki sürüklenmenin büyüklüğünün daha büyük olacağını öngörür. Sürüklenmenin büyüklüğü, seçilim katsayısı etkin popülasyon büyüklüğüne bölündüğünde 1'den az olduğunda herhangi bir alel frekansında seçilimi bastıracak kadar büyüktür. Bu nedenle, mutasyon ve genetik sürüklenmenin çarpımından kaynaklanan adaptif olmayan evrimin, öncelikle küçük, izole popülasyonlarda evrimsel değişimin sonuçsal bir mekanizması olduğu düşünülmektedir.[31] Genetik sürüklenmenin matematiği etkin popülasyon büyüklüğüne bağlıdır, ancak bunun bir popülasyondaki gerçek birey sayısıyla nasıl ilişkili olduğu açık değildir.[17] Seçilim altında olan diğer genlere genetik bağlantı, nötr bir alelin maruz kaldığı etkili popülasyon boyutunu azaltabilir. Daha yüksek rekombinasyon oranı ile bağlantı azalır ve bununla birlikte etkin popülasyon büyüklüğü üzerindeki bu yerel etki de azalır.[32][33] Bu etki, moleküler verilerde yerel rekombinasyon oranı ile genetik çeşitlilik arasında bir korelasyon[34] ve kodlamayan DNA bölgelerindeki gen yoğunluğu ile çeşitlilik arasında negatif korelasyon olarak görülebilir.[35] Seçilim altında olan diğer genlere bağlanma ile ilişkili stokastiklik, örnekleme hatası ile aynı değildir ve bazen genetik sürüklenmeden ayırt etmek için genetik otostop olarak bilinir.[17]
Düşük alel frekansı, alelleri rastgele şans eseri elenmeye karşı daha savunmasız hale getirir, hatta doğal seçilimin etkisini geçersiz kılar. Örneğin, dezavantajlı mutasyonlar genellikle popülasyon içinde hızla elenirken, yeni avantajlı mutasyonlar genetik sürüklenme yoluyla kayba karşı neredeyse nötr mutasyonlar kadar savunmasızdır. Avantajlı mutasyon için alel frekansı belirli bir eşiğe ulaşana kadar genetik sürüklenmenin hiçbir etkisi olmayacaktır.[30]
Popülasyon darboğazı
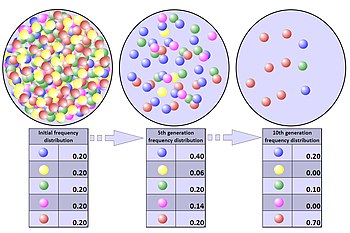
Popülasyon darboğazı, bir popülasyonun rastgele bir çevresel olay nedeniyle kısa bir süre içinde önemli ölçüde daha küçük bir boyuta inmesidir. Gerçek bir popülasyon darboğazında, popülasyonun herhangi bir üyesinin hayatta kalma olasılığı tamamen rastlantısaldır ve herhangi bir özel genetik avantajla iyileştirilmez. Darboğaz, seçilimden tamamen bağımsız olarak alel frekanslarında radikal değişikliklere yol açabilir.[36]
Bir popülasyon darboğazının etkisi, darboğaz doğal bir felaket gibi tek seferlik bir olaydan kaynaklansa bile devam edebilir. Olağandışı genetik dağılıma neden olan bir darboğazın ilginç bir örneği, Mikronezya'daki Pingelap atolünde toplam çubuk hücre renk körlüğüne (akromatopsi) sahip bireylerin nispeten yüksek oranıdır.[37] Bir darboğazdan sonra akraba evliliği artar. Bu durum, akrabalı yetiştirme depresyonu olarak bilinen bir süreçte, resesif zararlı mutasyonların verdiği zararı artırır. Bu mutasyonların en kötüleri seçilerek, genetik olarak bunlarla bağlantılı olan diğer alellerin arka plan seçilimi sürecinde kaybolmasına yol açar.[2] Çekinik zararlı mutasyonlar için bu seçilim, genetik arındırma nedeniyle darboğazın bir sonucu olarak artabilir. Bu da genetik çeşitliliğin daha da kaybolmasına yol açar. Buna ek olarak, popülasyon büyüklüğünde sürekli bir azalma, gelecek nesillerde sürüklenmeden kaynaklanan daha fazla alel dalgalanması olasılığını artırır.
Bir popülasyonun genetik çeşitliliği bir darboğaz nedeniyle büyük ölçüde azalabilir ve faydalı adaptasyonlar bile kalıcı olarak ortadan kalkabilir.[38] Varyasyon kaybı, hayatta kalan popülasyonu hastalık, iklim değişikliği veya mevcut gıda kaynağındaki değişim gibi yeni seçilim baskılarına karşı savunmasız bırakır, çünkü çevresel değişikliklere yanıt olarak uyum sağlamak, doğal seçilimin gerçekleşmesi için popülasyonda yeterli genetik varyasyon gerektirir.[39][40]
Yakın geçmişte bilinen birçok popülasyon darboğazı vakası yaşanmıştır. Avrupalıların gelişinden önce Kuzey Amerika çayırları milyonlarca büyük çayır tavuğunun yaşam alanıydı. Sadece Illinois'de sayıları 1900'de yaklaşık 100 milyon kuştan 1990'larda yaklaşık 50 kuşa düşmüştür. Nüfustaki düşüşler avlanma ve habitat tahribatından kaynaklandı, ancak bunun bir sonucu da türün genetik çeşitliliğinin çoğunun kaybolması oldu. Yüzyılın ortalarındaki kuşlarla 1990'lardaki kuşları karşılaştıran DNA analizi, sadece son birkaç on yılda genetik çeşitlilikte keskin bir düşüş olduğunu belgelemektedir. Şu anda büyük çayır tavuğu düşük üreme başarısı yaşamaktadır.[41]
Bununla birlikte, darboğaz ve genetik sürüklenmenin neden olduğu genetik kayıp, Ehrlichia'da olduğu gibi uygunluğu artırabilir.[42]
Aşırı avlanma, 19. yüzyılda kuzey deniz filinde de ciddi bir nüfus darboğazına neden olmuştur. Genetik çeşitlilikte ortaya çıkan düşüş, bu kadar agresif bir şekilde avlanmayan güney deniz filiinki ile karşılaştırılarak çıkarılabilir.[43]
Kurucu etkisi
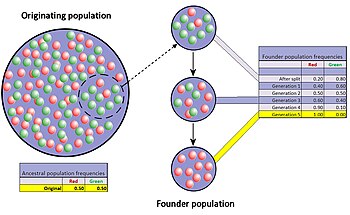
Kurucu etkisi, bir popülasyondaki küçük bir grup orijinal popülasyondan ayrılıp yeni bir popülasyon oluşturduğunda ortaya çıkan, popülasyon darboğazının özel bir durumudur. Yeni oluşan kolonideki rastgele alel örneklerinin, en azından bazı açılardan orijinal popülasyonu büyük ölçüde yanlış temsil etmesi beklenir.[44] Hatta orijinal popülasyondaki bazı genler için alel sayısının, kuruculardaki gen kopyası sayısından daha fazla olması ve tam temsili imkansız hale getirmesi bile mümkündür. Yeni oluşan bir koloni küçük olduğunda, kurucuları popülasyonun genetik yapısını gelecekte de güçlü bir şekilde etkileyebilir.
İyi belgelenmiş bir örnek, 1744 yılında Pensilvanya'ya yapılan Amiş göçünde bulunmaktadır. Yeni koloninin iki üyesi Ellis-Van Creveld sendromu için resesif aleli paylaşıyorlardı. Koloninin üyeleri ve onların soyundan gelenler dini izolatlar olma ve nispeten izole kalma eğilimindedirler. Nesiller boyu süren akraba evliliğinin bir sonucu olarak, Ellis-Van Creveld sendromu Amişler arasında genel nüfusa kıyasla çok daha yaygındır.[30][45]
Orijinal popülasyon ve koloni arasındaki gen frekanslarındaki fark, iki grubun birçok nesil boyunca önemli ölçüde farklılaşmasını da tetikleyebilir. Fark veya genetik mesafe arttıkça, ayrılan iki popülasyon hem genetik hem de fenetik olarak farklılaşabilir, ancak bu farklılaşmaya yalnızca genetik sürüklenme değil, doğal seçilim, gen akışı ve mutasyon da katkıda bulunur. Koloninin gen frekansındaki bu nispeten hızlı değişim potansiyeli, çoğu bilim insanının kurucu etkisini (ve buna bağlı olarak genetik sürüklenmeyi) yeni türlerin evriminde önemli bir itici güç olarak görmesine yol açmıştır. Sewall Wright, türleşmeye ilişkin değişen denge teorisiyle rastgele sürüklenmeye ve küçük, yeni izole edilmiş popülasyonlara bu önemi atfeden ilk kişi olmuştur.[46]
Wright'ın ardından Ernst Mayr, kurucu etkiyi takiben genetik varyasyondaki düşüşün ve küçük popülasyon boyutunun yeni türlerin gelişmesi için kritik öneme sahip olduğunu göstermek için birçok ikna edici model oluşturmuştur.[47] Bununla birlikte, hipotez deneysel araştırmalarla defalarca test edildiğinden ve sonuçlar en iyi ihtimalle belirsiz olduğundan, bugün bu görüş için çok daha az destek vardır.[48]
Tarihçe
Rastgele şansın evrimdeki rolü ilk olarak 1921 yılında Arend L. Hagedoorn ve A. C. Hagedoorn-Vorstheuvel La Brand tarafından özetlenmiştir.[49] Popülasyonlardaki varyasyon kaybında rastgele hayatta kalmanın önemli bir rol oynadığını vurgulamışlardır. Fisher (1922) buna, "Hagedoorn etkisi"nin ilk, ancak yanlış da olsa, matematiksel olarak ele alınmasıyla yanıt verdi.[50] Özellikle, sürüklenmenin etkilerinin önemli olması için birçok doğal popülasyonun çok büyük (N ~10.000) olduğunu ve sürüklenmenin evrimsel süreç üzerinde önemsiz bir etkiye sahip olacağını düşünmüştür. Düzeltilmiş matematiksel işlem ve "genetik sürüklenme" terimi daha sonra popülasyon genetiğinin kurucularından Sewall Wright tarafından ortaya atılmıştır. "Sürüklenme" terimini ilk kez 1929 yılında kullanmıştır, ancak o zamanlar bunu yönlendirilmiş bir değişim süreci veya doğal seçilim anlamında kullanıyordu.[51]
Örnekleme hatası yoluyla rastgele sürüklenme "Sewall-Wright etkisi" olarak bilinmeye başlandı, ancak Wright kendi adının verilmesinden hiçbir zaman tam olarak memnun olmadı. Wright, alel frekansındaki tüm değişiklikleri ya "sabit sürüklenme" (örn. seçilim) ya da "rastgele sürüklenme" (örn. örnekleme hatası) olarak adlandırmıştır.[52] "Sürüklenme" sadece stokastik anlamda teknik bir terim olarak benimsenmiştir.[53] Günümüzde genellikle örnekleme hatası açısından daha dar bir şekilde tanımlanmaktadır,[54] ancak bu dar tanım evrensel değildir.[55][56] Wright, ""rastgele sürüklenme" ya da hatta "sürüklenme "nin yalnızca bir bileşenle, örnekleme kazalarının etkileriyle sınırlandırılmasının kafa karışıklığına yol açma eğiliminde olduğunu" yazmıştır.[52] Sewall Wright, örnekleme hatası yoluyla rastgele genetik sürüklenme sürecini akraba evliliği yoluyla olana eşdeğer görmüştür, ancak daha sonraki çalışmalar bunların farklı olduğunu göstermiştir.[57]
Modern evrimsel sentezin ilk günlerinde, bilim insanları yeni popülasyon genetiği bilimini Charles Darwin'in doğal seçilim teorisiyle harmanlamaya başlamıştı. Bu çerçevede Wright, akraba evliliğinin nispeten izole küçük popülasyonlar üzerindeki etkilerine odaklandı. Küçük popülasyonlarda çapraz üreme ve genetik sürüklenme gibi olguların onları adaptif zirvelerden uzaklaştırabileceği ve bunun da doğal seçilimin onları yeni adaptif zirvelere doğru itmesine izin verebileceği adaptif bir manzara kavramını ortaya attı.[58] Wright, küçük popülasyonların doğal seçilim için daha uygun olduğunu düşünüyordu çünkü "akraba evliliği rastgele sürüklenme yoluyla yeni etkileşim sistemleri yaratmak için yeterince yoğundu ancak genlerin rastgele uyumsuz sabitlenmesine neden olacak kadar yoğun değildi".[59]
Wright'ın evrimsel şemada genetik sürüklenmenin rolü hakkındaki görüşleri neredeyse en başından beri tartışmalıydı. Wright'ı en yüksek sesle ve etkili biçimde eleştirenlerden biri meslektaşı Ronald Fisher'dı. Fisher, genetik sürüklenmenin evrimde bir rol oynadığını, ancak bunun önemsiz bir rol olduğunu düşünüyordu. Fisher, Wright'ın görüşlerini yanlış anlamakla suçlanmıştır çünkü eleştirilerinde Fisher, Wright'ın seçilimi neredeyse tamamen reddettiğini iddia eder gibi görünmüştür. Fisher'a göre, evrim sürecini uzun, istikrarlı, uyarlanabilir bir ilerleme olarak görmek, daha basit formlardan sürekli artan karmaşıklığı açıklamanın tek yoluydu. Ancak "aşamacılar" ile seçilim ve sürüklenmenin birlikte önemli bir rol oynadığı Wright evrim modeline daha sıcak bakanlar arasındaki tartışmalar devam etmiştir.[60]
1968 yılında Motoo Kimura, genetik değişikliklerin çoğunun nötr mutasyonlar üzerinde etkili olan genetik sürüklenmeden kaynaklandığını iddia eden nötral moleküler evrim teorisiyle tartışmayı yeniden alevlendirmiştir.[5][6]
Evrimde örnekleme hatası yoluyla genetik sürüklenmenin rolü, bağlantılı bölgelerdeki seçilimin daha önemli bir stokastik güç olduğunu savunan John H. Gillespie[61] ve William B. Provine tarafından eleştirilmiştir.
Ayrıca bakınız
Kaynakça
- ^ Gould SJ (2002). "Chapter 7, section "Synthesis as Hardening"". The Structure of Evolutionary Theory.
- ^ a b c Masel J (October 2011). "Genetic drift". Current Biology. 21 (20). Cell Press. ss. R837-8. doi:10.1016/j.cub.2011.08.007. PMID 22032182.
- ^ Star B, Spencer HG (May 2013). "Effects of genetic drift and gene flow on the selective maintenance of genetic variation". Genetics. 194 (1). ss. 235–44. doi:10.1534/genetics.113.149781. PMC 3632471 $2. PMID 23457235.
- ^ Miller 2000, s. 54
- ^ a b Kimura M (February 1968). "Evolutionary rate at the molecular level". Nature. 217 (5129). Nature Publishing Group. ss. 624–6. Bibcode:1968Natur.217..624K. doi:10.1038/217624a0. PMID 5637732.
- ^ a b Futuyma 1998, s. 320
- ^ Stoltzfus A (1999). "On the Possibility of Constructive Neutral Evolution". Journal of Molecular Evolution (İngilizce). 49 (2). ss. 169–181. Bibcode:1999JMolE..49..169S. doi:10.1007/PL00006540. ISSN 0022-2844. PMID 10441669. 30 Temmuz 2022 tarihinde kaynağından arşivlendi. Erişim tarihi: 20 Ocak 2022.
- ^ Muñoz-Gómez SA, Bilolikar G, Wideman JG, Geiler-Samerotte K (April 2021). "Constructive Neutral Evolution 20 Years Later". Journal of Molecular Evolution. 89 (3). ss. 172–182. Bibcode:2021JMolE..89..172M. doi:10.1007/s00239-021-09996-y. PMC 7982386 $2. PMID 33604782.
- ^ "Sampling Error and Evolution". Understanding Evolution. University of California, Berkeley. 8 Aralık 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 1 Aralık 2015.
- ^ Wahl LM (August 2011). "Fixation when N and s vary: classic approaches give elegant new results". Genetics. 188 (4). Genetics Society of America. ss. 783–5. doi:10.1534/genetics.111.131748. PMC 3176088 $2. PMID 21828279.
- ^ a b Hartl & Clark 2007, s. 112
- ^ Tian 2008, s. 11
- ^ Moran PA (1958). "Random processes in genetics". Mathematical Proceedings of the Cambridge Philosophical Society. 54 (1). ss. 60–71. Bibcode:1958PCPS...54...60M. doi:10.1017/S0305004100033193.
- ^ a b Charlesworth B (March 2009). "Fundamental concepts in genetics: effective population size and patterns of molecular evolution and variation". Nature Reviews. Genetics. 10 (3). Nature Publishing Group. ss. 195–205. doi:10.1038/nrg2526. PMID 19204717.
- ^ Der R, Epstein CL, Plotkin JB (September 2011). "Generalized population models and the nature of genetic drift". Theoretical Population Biology. 80 (2). Elsevier. ss. 80–99. doi:10.1016/j.tpb.2011.06.004. PMID 21718713.
- ^ Li & Graur 1991, s. 28
- ^ a b c Gillespie JH (November 2001). "Is the population size of a species relevant to its evolution?". Evolution; International Journal of Organic Evolution. 55 (11). John Wiley & Sons for the Society for the Study of Evolution. ss. 2161–9. doi:10.1111/j.0014-3820.2001.tb00732.x. PMID 11794777.
- ^ Neher RA, Shraiman BI (August 2011). "Genetic draft and quasi-neutrality in large facultatively sexual populations". Genetics. 188 (4). Genetics Society of America. ss. 975–96. arXiv:1108.1635 $2. doi:10.1534/genetics.111.128876. PMC 3176096 $2. PMID 21625002.
- ^ Ewens 2004
- ^ Li & Graur 1991, s. 29
- ^ Barton et al. 2007, s. 417
- ^ Futuyma 1998, s. 300
- ^ Otto SP, Whitlock MC (June 1997). "The probability of fixation in populations of changing size" (PDF). Genetics. 146 (2). Genetics Society of America. ss. 723–33. doi:10.1093/genetics/146.2.723. PMC 1208011 $2. PMID 9178020. 19 Mart 2015 tarihinde kaynağından (PDF) arşivlendi.
- ^ Cutter AD, Choi JY (August 2010). "Natural selection shapes nucleotide polymorphism across the genome of the nematode Caenorhabditis briggsae". Genome Research. 20 (8). Cold Spring Harbor Laboratory Press. ss. 1103–11. doi:10.1101/gr.104331.109. PMC 2909573 $2. PMID 20508143.
- ^ Hedrick 2005, s. 315
- ^ Li & Graur 1991, s. 33
- ^ Kimura & Ohta 1971
- ^ Masel J, King OD, Maughan H (January 2007). "The loss of adaptive plasticity during long periods of environmental stasis". The American Naturalist. 169 (1). University of Chicago Press on behalf of the American Society of Naturalists. ss. 38–46. doi:10.1086/510212. PMC 1766558 $2. PMID 17206583.
- ^ "Natural Selection: How Evolution Works". Actionbioscience. Washington, D.C.: American Institute of Biological Sciences. 6 Ocak 2010 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Kasım 2009. An interview with Douglas J. Futuyma. See answer to question: Is natural selection the only mechanism of evolution?
- ^ a b c Cavalli-Sforza, Menozzi & Piazza 1996
- ^ Zimmer 2001
- ^ Golding 1994, s. 46
- ^ Charlesworth B, Morgan MT, Charlesworth D (August 1993). "The effect of deleterious mutations on neutral molecular variation" (PDF). Genetics. 134 (4). Genetics Society of America. ss. 1289–303. doi:10.1093/genetics/134.4.1289. PMC 1205596 $2. PMID 8375663. 12 Mart 2020 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 9 Aralık 2015.
- ^ Presgraves DC (September 2005). "Recombination enhances protein adaptation in Drosophila melanogaster". Current Biology. 15 (18). Cell Press. ss. 1651–6. doi:10.1016/j.cub.2005.07.065. PMID 16169487.
- ^ Nordborg M, Hu TT, Ishino Y, Jhaveri J, Toomajian C, Zheng H, Bakker E, Calabrese P, Gladstone J, Goyal R, Jakobsson M, Kim S, Morozov Y, Padhukasahasram B, Plagnol V, Rosenberg NA, Shah C, Wall JD, Wang J, Zhao K, Kalbfleisch T, Schulz V, Kreitman M, Bergelson J (July 2005). "The pattern of polymorphism in Arabidopsis thaliana". PLOS Biology. 3 (7). Public Library of Science. ss. e196. doi:10.1371/journal.pbio.0030196. PMC 1135296 $2. PMID 15907155.

- ^ Robinson R, (Ed.) (2003). "Population Bottleneck". Genetics. 3. New York: Macmillan Reference USA. ISBN 0-02-865609-1. LCCN 2002003560. OCLC 614996575. Erişim tarihi: 14 Aralık 2015.
- ^ Hussels IE, Morton NE (May 1972). "Pingelap and Mokil Atolls: achromatopsia". American Journal of Human Genetics. 24 (3). ss. 304–309. PMC 1762260 $2. PMID 4555088.
- ^ Futuyma 1998, ss. 303–304
- ^ O'Corry-Crowe G (March 2008). "Climate change and the molecular ecology of Arctic marine mammals". Ecological Applications. 18 (2 Suppl). Ecological Society of America. ss. S56-76. doi:10.1890/06-0795.1. PMID 18494363.
- ^ Cornuet JM, Luikart G (December 1996). "Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data". Genetics. 144 (4). Genetics Society of America. ss. 2001–14. doi:10.1093/genetics/144.4.2001. PMC 1207747 $2. PMID 8978083.
- ^ Sadava et al. 2008, chpts. 1, 21–33, 52–57
- ^ Dale C, Moran NA (August 2006). "Molecular interactions between bacterial symbionts and their hosts". Cell. 126 (3). ss. 453–65. doi:10.1016/j.cell.2006.07.014. PMID 16901780.
- ^ "Bottlenecks and founder effects". Understanding Evolution. University of California, Berkeley. 4 Aralık 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 14 Aralık 2015.
- ^ Campbell 1996, s. 423
- ^ "Genetic Drift and the Founder Effect". Evolution Library (Web resource). Evolution. Boston, MA: WGBH Educational Foundation; Clear Blue Sky Productions, Inc. 2001. OCLC 48165595. 14 Mart 2009 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Nisan 2009.
- ^ Wolf, Brodie & Wade 2000
- ^ Hey, Fitch & Ayala 2005
- ^ Howard & Berlocher 1998
- ^ Hagedoorn AL, Hagedoorn-Vorstheuvel La Brand AC (1921). The Relative Value of the Processes Causing Evolution. The Hague: Martinus Nijhoff. 28 Mart 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 28 Mart 2019.
- ^ Fisher RA (1922). "On the Dominance Ratio". Proceedings of the Royal Society of Edinburgh. Cilt 42. ss. 321–341. doi:10.1017/s0370164600023993.
- ^ Wright S (November–December 1929). "The evolution of dominance". The American Naturalist. 63 (689). Chicago, IL: University of Chicago Press on behalf of the American Society of Naturalists. ss. 556–561. doi:10.1086/280290. ISSN 0003-0147. JSTOR 2456825.
- ^ a b Wright S (1955). "Classification of the factors of evolution". Cold Spring Harbor Symposia on Quantitative Biology. Cilt 20. Cold Spring Harbor Laboratory Press. ss. 16–24D. doi:10.1101/SQB.1955.020.01.004. PMID 13433551. Symposium: "Population Genetics: The Nature and Causes of Genetic Variability in Populations".
- ^ Stevenson 1991
- ^ Freeman & Herron 2007
- ^ Masel J (August 2012). "Rethinking Hardy-Weinberg and genetic drift in undergraduate biology". BioEssays. 34 (8). John Wiley & Sons. ss. 701–710. doi:10.1002/bies.201100178. PMID 22576789.
- ^ Lynch 2007
- ^ Crow JF (March 2010). "Wright and Fisher on inbreeding and random drift". Genetics. 184 (3). Genetics Society of America. ss. 609–611. doi:10.1534/genetics.109.110023. PMC 2845331 $2. PMID 20332416.
- ^ Larson 2004, ss. 221–243
- ^ Stevenson 1991: Quote attributed to William B. Provine in The Origins of Theoretical Population Genetics (1971), p. 162; Chicago: University of Chicago Press.
- ^ Avers 1989
- ^ Gillespie JH (June 2000). "Genetic drift in an infinite population. The pseudohitchhiking model". Genetics. 155 (2). Genetics Society of America. ss. 909–919. doi:10.1093/genetics/155.2.909. PMC 1461093 $2. PMID 10835409.
Konuyla ilgili yayınlar
- Avers, Charlotte J. (1989). Process and Pattern in Evolution. New York: Oxford University Press. ISBN 0-19-505275-7. LCCN 88005368. OCLC 17677554.
- Barton, Nicholas H.; Briggs, Derek E.G.; Eisen, Jonathan A.; Goldstein, David B.; Patel, Nipam H. (2007). Evolution. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. ISBN 978-0-87969-684-9. LCCN 2007010767. OCLC 86090399.
- Campbell, Neil A. (1996). Biology. 4th. Menlo Park, CA: Benjamin/Cummings Pub. Co. ISBN 0-8053-1940-9. LCCN 95045572. OCLC 33333455.
- Cavalli-Sforza, L. Luca; Menozzi, Paolo; Piazza, Alberto (1996). The History and Geography of Human Genes. Abridged paperback. Princeton, N.J.: Princeton University Press. ISBN 0-691-02905-9. OCLC 35527063.
- Ewens, Warren J. (2004). Mathematical Population Genetics I. Theoretical Introduction. 2nd. 27. New York: Springer-Verlag. ISBN 0-387-20191-2. LCCN 2003065728. OCLC 53231891.
- Freeman, Scott; Herron, Jon C. (2007). Evolutionary Analysis. 4th. Upper Saddle River, NJ: Pearson Prentice Hall. ISBN 978-0-13-227584-2. LCCN 2006034384. OCLC 73502978.
- Futuyma, Douglas (1998). Evolutionary Biology. 3rd. Sunderland, MA: Sinauer Associates. ISBN 0-87893-189-9. LCCN 97037947. OCLC 37560100.
- Golding, Brian, (Ed.) (1994). Non-Neutral Evolution: Theories and Molecular Data. New York: Chapman & Hall. ISBN 0-412-05391-8. LCCN 93047006. OCLC 29638235. "Papers from a workshop sponsored by the Canadian Institute for Advanced Research."
- Hartl, Daniel L.; Clark, Andrew G. (2007). Principles of Population Genetics. 4th. Sunderland, MA: Sinauer Associates. ISBN 978-0-87893-308-2. LCCN 2006036153. OCLC 75087956.
- Hedrick, Philip W. (2005). Genetics of Populations. 3rd. Boston, MA: Jones and Bartlett Publishers. ISBN 0-7637-4772-6. LCCN 2004056666. OCLC 56194719.
- Hey, Jody; Fitch, Walter M.; Ayala, Francisco J., (Ed.) (2005). Systematics and the Origin of Species: On Ernst Mayr's 100th Anniversary. Washington, D.C.: National Academies Press. ISBN 978-0-309-09536-5. LCCN 2005017917. OCLC 70745851.
- Howard, Daniel J.; Berlocher, Steward H., (Ed.) (1998). Endless Forms: Species and Speciation. New York: Oxford University Press. ISBN 978-0-19-510901-6. LCCN 97031461. OCLC 37545522.
- Kimura, Motoo; Ohta, Tomoko (1971). Theoretical Aspects of Population Genetics. Monographs in Population Biology. 4. Princeton, NJ: Princeton University Press. ss. 1–219. ISBN 0-691-08096-8. LCCN 75155963. OCLC 299867647. PMID 5162676.
- Larson, Edward J. (2004). Evolution: The Remarkable History of a Scientific Theory. Modern Library Chronicles. 17. New York: Modern Library. ISBN 0-679-64288-9. LCCN 2003064888. OCLC 53483597.
- Li, Wen-Hsiung; Graur, Dan (1991). Fundamentals of Molecular Evolution. Sunderland, MA: Sinauer Associates. ISBN 0-87893-452-9. LCCN 90043581. OCLC 22113526.
- Lynch, Michael (2007). The Origins of Genome Architecture. Sunderland, MA: Sinauer Associates. ISBN 978-0-87893-484-3. LCCN 2007000012. OCLC 77574049.
- Miller, Geoffrey (2000). The Mating Mind: How Sexual Choice Shaped the Evolution of Human Nature. New York: Doubleday. ISBN 0-385-49516-1. LCCN 00022673. OCLC 43648482.
- Sadava, David; Heller, H. Craig; Orians, Gordon H.; Purves, William K.; Hillis, David M. (2008). Life: The Science of Biology. 8th. II: Evolution, Diversity and Ecology. Sunderland, MA; Gordonsville, VA: Sinauer Associates; W. H. Freeman and Company. ISBN 978-0-7167-7674-1. LCCN 2006031320. OCLC 71632224.
- Stevenson, Joan C. (1991). Dictionary of Concepts in Physical Anthropology. Reference Sources for the Social Sciences and Humanities. 10. Westport, CT: Greenwood Press. ISBN 0-313-24756-0. LCCN 90022815. OCLC 22732327.
- Tian, Jianjun Paul (2008). Evolution Algebras and their Applications. Lecture Notes in Mathematics. 1921. Berlin; New York: Springer. doi:10.1007/978-3-540-74284-5. ISBN 978-3-540-74283-8. LCCN 2007933498. OCLC 173807298. Zbl 1136.17001.
- Wolf, Jason B.; Brodie, Edmund D.; Wade, Michael J., (Ed.) (2000). Epistasis and the Evolutionary Process. Oxford, UK; New York: Oxford University Press. ISBN 0-19-512806-0. LCCN 99046515. OCLC 42603105.
- Zimmer, Carl (2001). Evolution: The Triumph of an Idea. 1st. Introduction by Stephen Jay Gould; foreword by Richard Hutton. New York: HarperCollins. ISBN 0-06-019906-7. LCCN 2001024077. OCLC 46359440.
Dış bağlantılar
- Sheehy, Bob. "Population genetics simulation program". Radford, VA: Radford University. Erişim tarihi: 21 Aralık 2015. Arşivlenmesi gereken bağlantıya sahip kaynak şablonu içeren maddeler (link)
- Grimes, Bill. "Genetic Drift Simulation". Tucson, Arizona: The University of Arizona. Erişim tarihi: 25 Ağustos 2016. Arşivlenmesi gereken bağlantıya sahip kaynak şablonu içeren maddeler (link)










